






CONTINUACIÓN CLASE: COMPLEJO MAYOR DE HISTOCOMPATIBILIDAD

Genes
del MHC que se encuentra en todos los animales, también los humanos. En los
humanos esos cromosomas están en el brazo corto del cromosoma 6 y esos genes se
pueden clasificar en dos clases, genes de clase 1 y genes de clase 2. Hay muchos
genes de MHC, uno de estos genes se llaman genes clásicos que codifican para un
tipo de proteínas que tienen la función de presentar péptidos a los LT. Los
genes clásicos de clase 1 (A, B, C) van a codificar unas proteínas que están
presentes en todas las células nucleadas y a través de esa molécula se va hacer
presentación cuando la célula este infectada por un microorganismo o cuando se está
desarrollando como una célula cancerosa o tumoral.
La
mejor defensa en el momento de que una célula esta infectada o sea tumoral es
la apoptosis. Las mejores células para destruir esas células infectadas o
tumorales son los LTCD8.
La
forma de alertar de que la célula está infectada o desarrollando cáncer es a
través de las moléculas de MHC clase I.
Los
otros tipos de genes clásicos de clase 2 que se llaman DP, DQ, DR, codifican
para unas moléculas que tienen la función de presentar péptidos a los LTCD4. La
presentación la hace una célula a través de las moléculas del MHC.
Todas
las células poseen moléculas de MHC clase I, porque todas son susceptibles de
ser infectadas o de desarrollar cáncer. Las Células presentadoras de antígenos
expresan moléculas de MHC clase II (también células endoteliales, células
epiteliales timicas pueden hacer presentación, pero no son presentadoras
profesionales).
12
moléculas distintas de MHC por individuo. Con 12 moléculas distintas un
individuo puede presenta hasta 10 millones de péptidos distintos porque cada molécula
es capaz de reconocer muchos péptidos diferentes pero solo puede presentar un
péptido a la vez.
La
diferencia entre cada alelo está concentrada en la forma de hendidura, la forma
de la hendidura determina los péptidos que se pueden unir para ser presentados.
Todos
poseemos los mismos genes del HLA, la diferencia radica en los alelos. Es por
esto que cada uno de nosotros aunque tengamos las mismas moléculas, presenta
diferentes péptidos.
¿La
Diversidad (a nivel poblacional/polimorfismo) es buena o mala? La importancia
de la diversidad es que no todos a nivel poblacional van a poder generar una
respuesta frente a los mismos microorganismos, hay personas que se infectan y
que pueden que generar una respuesta en contra de los microorganismos y esos
son aquellos que pudieron tomar antígenos del microorganismo y presentarlo a
los LT. Hay otras personas que por la misma diversidad no son capaces de
presentar los péptidos de las demás personas, aquellas personas que no puedan
presentar péptidos derivados de un microorganismo, van a padecer la enfermedad.
Se infectan tanto los que se pueden defender como los que no se pueden
defender, se enferman los que no pueden presentar péptidos. El concepto de la
diversidad es importante porque no hace que todos seamos susceptibles a padecer
las mismas enfermedades, porque hay individuos capaces de presentar péptidos y
de generar una respuesta inmune frente a esos microorganismos pero hay otros
que son incapaces.
Diversidad:
Determina la capacidad generar una respuesta inmune en contra de un antígeno.
Ser inmune o susceptible, susceptible significa que se infecta pero no es capaz
de defender frente a ese riesgo, capacidad de presentar o no péptidos derivados
de los microorganismos. Esto explica el porqué el ser vacunado y no presentar
péptidos del microorganismo.
La
importancia de la diversidad es que a nivel poblacional estamos protegidos como
especie.
Pero la diversidad tiene también desventajas:
·
Lo que
determina que un trasplante se acepte o no es la similitud entre los alelos del
HLA, como alelicamente somos diferentes, la probabilidad de encontrar un
donante y un receptor compatible es muy bajo, la diversidad por lo tanto limita
el uso de los trasplantes como una herramienta para tratar diferentes
condiciones clínicas. Similitud entre alelos del donante y receptor.
·
No el 100% de
la población que reciben una vacuna no generan memoria, ya que por la misma
diversidad, no todos presentamos los péptidos derivados de las vacunas.
En
el contexto general la diversidad es buena, pero esa misma diversidad limita la
presentación antigénica de determinados epitopes de un antígeno
Características
que determinan la diversidad
Poligenia:
Distintos genes que codifican para unas proteínas similares en estructura y
función, pero cada gen (alelo) va a producir moléculas con hendiduras
diferentes.
Duplicación de
los genes DR: Aumenta el número de
moléculas del MHC, presentara mayor número de genes. Cuando se van a hacer
trasplantes, este es el alelo que más se va a tipificar ya que si nos parecemos
en DRb
la probabilidad de que nos parezcamos en los otros genes es más alta. Al
aparecer genes extra del MHC se va a aumentar el número de moléculas, esa
duplicación permite generar hasta 18 moléculas en aquellas personas que tengan
duplicado los genes en los dos alelos.
Dentro
del mismo brazo corto del cromosoma 6 aparecen otros genes del MHC que se
denominan genes no clásicos, de estos genes se van a producir algunas proteínas
que no presentan péptidos a los LT pero que pueden ayudar en la presentación.
Los genes no clásicos del MHC se van a encontrar proteínas que no presentan
péptidos pero que ayudan a que se haga la presentación. Encontramos genes no
clásicos de clase I y genes no clásicos de clase II.
Hay
genes no clásicos de clase I que son
el HLA-G, HLA-E, HLA-F. Estos genes codifican para moléculas del MHC de clase
I. Con estas proteínas no se presentan péptidos a los LTCD8. La importancia de
estas moléculas no clásicas de clase I es que a través de estas moléculas es
que se inhibe la acción citotóxicas NK sobre células propias. Las células NK
tienen la capacidad de destruir células propias que estén infectadas o
tumorales, pero hay que inhibirlas porque siempre están en un estado de querer
destruir. Las moléculas no clásicas de clase I se convierten en moléculas que
inhiben a las células NK, y esto es importante porque así las células NK no van
a destruir células propias. Hay una condición clínica que se ve en el HLA-G que
se llama abortos espontáneos recurrentes, en estas mujeres hay fecundación
normal, implantación normal, pero cuando empieza la placentación las células NK
atacan a la placenta y esto es debido a una ausencia de las moléculas del HLA-G,
también se ha visto por deficiencia de moléculas de HLA-E.
Se
dice que el embarazo es un periodo de inmunosupresión, pero esto es falso
porque el embarazo es un periodo de inmunomodulación que consiste en cambiar la
manera de responder. Las mujeres generalmente presentan respuestas
proinflamatorias que son muy buenas reaccionando frente a las infecciones
virales y potencian muchos macrófagos, por lo que las mujeres reaccionan mejor
a las infecciones virales que los hombres. Cuando una mujer entra en embarazo
cambia la respuesta proinflamatoria a una respuesta más antiinflamatoria (inmunomodulación),
en una mujer gestante se busca que no active tantas celulas NK, LTCD8 ni
macrófago, sino mas bien respuesta antiinflamatoria mas mediada por celulas
como Eosinofilos, Basofilos, es por esto que a la mujer gestante le de tanta
gripa y poco frecuente las infecciones parasitarias porque la respuesta
antiinflamatoria (Th2) es muy buena contra los parásitos. Cambio en el enfoque
de la respuesta inmune.
Las
moléculas no clásicas de clase I inhiben a las células NK para que destruyan células
propias.
Genes no
clásicos de clase II, estos genes si
codifican para unas proteínas que van a ayudar a que se haga la presentación.
No van a codificar proteínas para presentar pero si van a codificar proteínas
para la presentación. Estos genes van a codificar para unas proteínas que van a
“Ayudar” a presentar, unas ayudan a degradar los péptidos, otras ayudan a que
el péptido se una a la molécula del MHC.
Dentro
del mismo brazo corto del cromosoma 6 va a aparecer un tercer grupo de genes que
se va a llamar genes del MHC clase III, estos genes no tienen nada que ver ni
con procesamiento ni con presentación de antígeno. Los genes de clase III
codifican para proteínas del sistema del complemento y para algunas citoquinas.
Dos
factores están asociados con la diversidad, los cuales son la raza y la
geografía.
Algunos alelos están más frecuentes en ciertas
poblaciones que en otras poblaciones. La diversidad alelica tiene una fuerte
asociación racial, por lo tanto entre más cercano estemos racialmente, mayor
probabilidad de que nos parezcamos alelicamente. Este es un concepto que se
aplica muy frecuente en los trasplantes. Aunque ese concepto de raza se ha distorsionado
por la globalización, porque socialmente nos hemos recombinado.
5
factores por los que somos diversos:
- Poligenia
- Duplicación génica
- Polimorfismo: Una variación superior al 1% de alelos en la población. Muchos genes tienen varios alelos pero por tener varios alelos no significan que sean polimórficos. Se considera polimórfico cuando su diversidad o variación supere el 1% de la población.
No
todos los alelos codifican para proteínas, en realidad los alelos que codifican
para proteínas es muy reducido. Hay alelos donde la diversidad es mayor,
mientras se encuentran alelos mucho mas conservados en la población. Si todos
somos distintos alelicamente.
¿Por
qué si se pueden realizar trasplantes? Poblacionalmente no hay dos personas con
los mismos alelos a no ser que sean hermanos gemelos homogocitos, pero hay
personas que comparten ciertos alelos. La idea de que no somos idénticos es que
dos individuos no tienen los mismos alelos para cada gen pero en la población
si compartimos ciertos alelos, y esto es lo que permite plantear la opción de los
trasplantes.
No
somos idénticos en todos los alelos, pero si compartimos determinados alelos a
nivel poblacional. Entre más cercanos estemos racialmente y geográficamente hay
mayor probabilidad de compartir determinados números de alelos.
Hay
trasplantes en los que la homología es mayor o menor, un trasplante en el que
la homología no se necesita que sea muy alta, homología entre donante y
receptor no tiene que ser muy alta, con que sea incluso solo homologo en el 40%
vale. Hay unos en cambio que necesitan homología del 97% como en el caso de
medula ósea, es por esto que el trasplante de medula ósea generalmente proviene
de hermanos donde la probabilidad es mucho mayor.
¿Cómo
se llama la prueba para comprobar la homología alelica? Se llama tipificación
de alelos del HLA. La tipificación es una prueba para conocer los alelos, es
una reacción en cadena de la polimerasa (PCR). El gen que siempre se va a
tipificar es el DR porque es el más polimórfico y si se asume que dos personas
comparten el alelo Beta del gen DR, la probabilidad de que compartan otros
alelos es mayor.
De
cada alelo sale una molécula con una hendidura diferente a otro alelo.
El
polimorfismo va a determinar que péptidos van a ser presentados por cada
individuo. Lo único distinto entre las moléculas derivadas de diferentes alelos
es la forma de la hendidura y es por esto cada alelo va a codificar para una
proteína que presenta péptidos distintos, esto aplica tanto para moléculas de
clase I como para moléculas de clase II.
- Otro factor que aumenta la diversidad se llama Recombinación de haplotipos.
Ningún
hijo va a ser idéntico a ninguno de los dos padres, porque la idea es que cada
hijo tenga la mitad de uno de sus padres. (Jhon y Andrew tienen los mismos
alelos porque en la división meiotica de cada padre se dio igual). Hay personas
que no tiene expresado todos sus alelos porque tienen otro que cumple la misma
función.
Segregación
mendeliana, los hijos van a tener una característica del padre y una
característica de la madre.
Haplotipo son los
alelos sobre un cromosoma. El padre
tiene haplotipo 1 y haplotipo 2, la madre tiene haplotipo 1 y haplotipo 2. Los
hijos heredan el haplotipo 1 o 2 de cada padre. Jaime heredo haplotipo 2 del
padre y haplotipo 2 de la madre. Cada hijo hereda un haplotipo o del papa o de
la mama. Jaime tiene un haplotipo nuevo que heredo de los padres, presenta
péptidos distintos a los de sus hermanos.
La
recombinación de haplotipos aumenta el número de moléculas en la población, va
a aportar para la diversidad porque va a dar origen a nuevos haplotipos que son
distintos a los padres.
2. Otra
condición que cuenta para la diversidad se llama la expresión codominante
La
mayoría de genes se expresan dominantemente. Por ejemplo tenemos un gen que
determina el color de ojos pero se tiene un color dado por la madre (alelo) y
otro color dado por el papa (alelo), es decir, de cada gen se tiene dos alelos
y se expresa uno de ellos, el dominante. Al hijo se le puede heredar o el alelo
dominante o el recesivo. En caso de los genes del HLA se expresan
codominantemente, esto significa que en una misma célula se van a expresar
tanto los alelos que provienen del padre como los alelos que provienen de la
madre. Lo que se logra gracias a la expresión codominante es que se aumenta el
número de moléculas expresadas al mismo tiempo y al aumentarse el número de
moléculas expresadas se aumenta la cantidad de péptidos que se pueden
presentar.
Si
los genes del HLA se expresaran dominantemente las moléculas que se pudieran
expresar al mismo tiempo seria 6, del papa o de la mama. Pero a tener una
expresión codominante se pueden expresar todas las moléculas al mismo tiempo.
Nomenclatura:
Nomenclatura:
HLA
–DRB1*0401
- HLA: MHC en humanos.
- D: Clase II
- R: Familia, que pueden ser P, Q ó R.
- B: Del gen Beta, porque los genes de clase II tienen gen Alfa y gen Beta.
- B1: Porque del DR hay B1, B3, B4 y B5 cuando hay duplicación.
*0401:
Lo que viene después del *
es el alelo
·
Los genes de
clase I serian o A, B o C y después viene el alelo. En los genes de clase I no
hay ni Alfa ni Beta, porque los genes de clase 1 codifican para una sola cadena
que es la cadena Alfa.
Hay
una fuerte asociación entre la expresión de ciertos alelos con determinadas
enfermedades. Hay muchas condiciones y sobre todo enfermedades autoinmunes que
se han relacionado con la expresión de ciertos alelos, esto significa que tener
un determinado alelo predispone a padecer la enfermedad autoinmune. El ejemplo
mas claro de esto es la Espondilitis Anquilosante una enfermedad donde hay
fusión de vertebras, la asociación de esta enfermedad se encuentra en la
expresión del alelo HLA-B27 (presente hasta en el 97% de las personas con la
enfermedad, pero no del 100% porque hay personas que tienen el alelo pero no presentan
la enfermedad). Otra enfermedad es el Lupus Eritematoso Sistémico asociada con
el alelo HLA-DR3. Por lo tanto ciertas enfermedades autoinmunes están
condicionadas a la presencia de determinados alelos, pero esto no se tiene muy
claro.
Procesamiento y
Presentación de antígenos
Hay
una célula capaz de reconocer el microorganismo, internalizar el
microorganismo, destruir el microorganismo para a través de moléculas del MHC
presentar fragmentos de ese microorganismo, esa presentación la va hacer a los LT.
Como en este caso se está hablando de un microorganismo que esta presenta
extracelularmente la célula que reconoce el microorganismo seria una APC por lo
que el le presenta el fragmento al LTD4. A esto se la llamaría vía de
procesamiento y presentación de clase II.
Pero
si habláramos de una célula tumoral o una célula infectada con un
microorganismo, ese microorganismo va a ser destruido y a través de moléculas
del MHC de clase I se va a presentar un péptido a un LTCD8. A esto se le llama vía
de procesamiento y presentación de clase I.
La
idea general es que los microorganismos son reconocidos o infectan una célula.
La célula que lo reconoce o la célula infectada pueden destruir o degradar el
microorganismo o las proteínas del microorganismo para posteriormente unirlas a
las moléculas del MHC para hacerle la presentación a los LT, LTCD4 o LTCD8 de acuerdo al tipo u origen del antígeno.
Vía de
Procesamiento de presentación de clase II
Esta vía la van a realizar las células
presentadoras de antígeno (APC), sobre todo las APC profesionales que son: las células
dendríticas, los LB o los macrófagos. En este caso estas células APC tienen o
expresan receptores que le permitan reconocer los microorganismos, después los
internaliza, los degrada y se los presenta a los LTCD4.
Las mejores APC son las células dendríticas
por varias características:
- Estas células están ubicadas en los sitios de entrada o en los principales sitios de entrada de los microorganismos como lo son los epitelios
- Tienen una morfología donde proyectan muchas dendritas y con esto pueden captar muchos microorganismos o antígenos.
- Estas células dendríticas sufren algo que se llama maduración. La maduración se da cuando una célula dendrítica de un epitelio reconoce un microorganismo se dice que está en un estado inmaduro o inactivo, pero el reconocimiento de ese microorganismo le induce migrar hacia un órgano linfoide secundario y en este viaje desde el sitio de la infección al lugar de presentación esa célula se activa y así madura.
¿Qué cambios se dan entre una célula
dendrítica inactiva a una célula dendrítica activa?
Las células dendríticas en los epitelios
expresan muchos receptores PRRs (receptores para PAMPs o DAMPs). Al expresar
muchos PRRs, en los epitelios, tienen una función principal que es la de
reconocer pero cuando reconocen y comienzan a migrar cambia, deja de expresar
estos receptores y aumenta la expresión de moléculas del MHC y también aumenta
la expresión de un grupo de moléculas que se llaman moléculas coestimuladoras (moléculas
B7)

Como células activadas ya su función no es
reconocer, su función es presentar. Inactiva es muy buena reconociendo,
activada es muy buena presentando.
Una célula dendrítica activada puede expresar
alrededor de 7 millones de moleculas del MHC. Esas B7 favorecen que se den la
presentación.
¿Cómo es esta vía de procesamiento y
presentación?
La célula dendrítica, macrófago o LB, a través de PRRs reconocen o microorganismo o
moléculas derivadas de estos microorganismos, los internaliza. Una vez
internalizados se forma una vesícula que se llama endosoma o fagosoma, esta
vesícula se une con otra vesícula que se llama lisosoma, los lisosomas
contienen lisosimas que son un tipo de enzimas (proteasas, lipasas, DNAasas,
RNAasas, fosfatasas), estas lisosimas que están en el lisosoma están inactivas.
Cuando se fusionan las dos vesículas (fagosoma y lisosoma) se forma una nueva
vesícula que se llama endolisosoma o fagolisosoma, cuando se forma el
endolisosoma o fagolisosoma hay un evento muy importante y es que al interior
de esta vesícula se cambia el pH y el pH baja, y cuando el pH baja se activa
las lisosimas. Las lisosimas activas degradan el microorganismo o degradan el
antígeno o degradan la molécula, y esa degradación va a dar origen a los
péptidos que van a ser presentados. Por lo tanto, la célula reconoce,
internaliza y degrada gracias a las lisosimas pero la actividad de las
lisosimas depende del cambio en el pH.
Mientras ocurre esa degradación, en el
retículo endoplasmatico se están sintetizando las moléculas de MHC de clase II.
Se sintetiza la cadena alfa y la cadena Beta, y mientras no tengan un péptido
estas moléculas son muy inestables, para estabilizar estas moleculas aparecen
unas acompañantes que se llaman Chaperonas y vamos hablar de dos. Una primer chaperona
se llama CALNEXINA, esta estabiliza la síntesis de las moleculas de MHC de
clase II y la segunda chaperona se llama la CADENA INVARIANTE, esta cadena
invariante también ayuda a esa estabilización. Cuando la molécula de MHC ya se
encuentra sintetizada y estabilizada, esta molecula es transportada por
vesículas, primero pasando por el aparato de golgi y después por el citoplasma
hasta llegar al lugar donde este el péptido el cual se encuentra en el
endolisosoma o fagolisosoma. La molécula MHC de clase II llega a la vesícula
pero con la cadena invariante que está ocupando la hendidura por lo que se debe
quitar la cadena invariante, para quitar la cadena invariante se van a dar dos
pasos:
- Las lisosimas que degradaron al microorganismo, degradan parte de la cadena invariante, quitan una parte pero queda otra parte. El fragmento de la cadena invariante que queda unido a la molécula de MHC de clase II se llama el CLIP. El clip queda ocupando justo la hendidura.
- El CLIP de la hendidura lo quita una molécula que se llama HLA-DM que viene de un gen no clásico.
Cuando se quita el CLIP la hendidura queda
libre y ahora si se van a unir los péptidos que se generaron por la degradación
del antígeno o del microorganismo. Cuando ya se tiene la molécula de MHC de
clase II más el péptido, se transporta hacia la membrana para presentarlo a un
LTCD4.

Los antígenos se degradan por lo tanto en el
endolisosoma o fagolisosoma, lo degrada las lisosimas. Las Catepsinas son un
grupo importante de lisosimas. La molécula de MHC de clase II se sintetiza en
el Reticulo endoplasmatico, las que participan en la síntesis son las
chaperonas las cuales son calnexina y cadena invariante. Cuando ya se forma la
molécula de MHC clase II viaja por vesículas y llega al endolisosoma o
fagolisosoma. Hay que quitar la cadena invariante, en un primer paso lo hace
las mismas lisosimas pero dejan el CLIP el cual está en la hendidura el cual lo
quita la HLA-DM que es una molécula no clásica. Cuando se quita el clip se
puede unir el péptido y ya se tiene la molécula estable para ir a presentar a
un LTCD4.
El + en el LTCD4+ indica que esa molécula
esta expresada

La
imagen anterior muestra la cadena invariante dentro del endolisosoma, las
lisosimas degradan parte de esa cadena invariante pero queda el CLIP que es
retirado por HLA-DM.
A
esta vesícula cuando contiene las moléculas de clase II, el HLA-DM y el
péptido, se puede llamar una vesícula o un compartimiento de clase II del MHC.
Vía de Procesamiento
de presentación de clase I
La
vía de la clase II la hacen células APC que reconocen extracelularmente, esta
vía de clase I por lo contrario la hace las células que estén infectadas o
tumorales.
Lo
que buscan los microorganismos al interior de la célula es replicarse y en esa
replicación se originan proteínas de los microorganismos. El ejemplo del que se
hablara es de una célula infectada por un virus. El virus infecta la célula y
se replica al interior de la célula, en ese tiempo de replicación se generan
proteínas virales.
Al
interior de todas nuestras células hay una proteína bien importante que se
llama la UBICUITINA cuya función es marcar proteínas que van a ser degradadas,
y es una proteína que funciona todo el tiempo porque van a marcar proteínas mal
plegadas, proteínas que ya cumplieron una función o en este caso proteínas
virales o proteínas tumorales.
La
Ubicuitina ubicuitiniza y las proteínas ubicuitinizadas son dirigidas a una
organela celular que se llama el proteosoma. La ubicuitina se le pega a las
proteínas y las lleva al proteosoma. El proteosoma cuenta con unas enzimas y
esas enzimas degradan las proteínas hasta originar péptidos, mientras esto está
ocurriendo se está sintetizando las moléculas del MHC clase I en el Retículo
Endoplasmatico.
En
el retículo endoplasmatico se tiene la cadena alfa del MHC de clase I y la b2m
(beta 2 microglobulina), cuando están separadas son inestables por lo que
tienen que aparecer chaperonas. Aparece una primera chaperona para la cadena
alfa que es la CALNEXINA (la misma de clase II). La b2m
no es una molécula del MHC porque no esta en estos genes. La b2m
es estabilizada por otras chaperonas, una es la TAPASINA, también la
CALRETICULINA y la molécula ERp57.
Cuando
ya se une la cadena alfa con la cadena b2m
desaparece la Calnexina. Aquí entra a jugar un papel importante la Tapasina la
cual va a acercar la molecula del MHC de clase I a otras proteínas que se
llaman las proteínas TAP. Hay dos tipos de proteínas TAP que se llaman: TAP1 y
TAP2 las cuales son proteínas que vienen de genes no clásicos del MHC. Las
proteínas TAP son las que transportan péptidos del citoplasma al retículo endoplasmatico.
El proteosoma degrado la proteína y origino los péptidos que quedaron en el
citoplasma, ahora se debe mover – traslocar los péptidos del citoplasma al
retículo endoplasmatico y esto lo hace las proteínas TAP, como la tapasina
acerco la molécula a TAP se le une fácilmente.
¿Cuál
es el tamaño de péptidos que se le puede unir a la hendidura de una molécula de
clase I del MHC? El proteosoma puede originar péptidos de hasta 16 aminoácidos
por lo que al retículo endoplasmatico pueden entrar un péptido que supere el
tamaño de la hendidura. El péptido hay que presentarlo y aquí participa ERAP,
ERAP es una aminopeptidasa del retículo endoplasmatico cuya función es que si
entran péptidos al retículo endoplasmatico de más de 11 aminoácidos ella los
corta para generar péptidos que ahora si tengan tamaño adecuado para unirse a
la molécula de MHC de clase I. Cuando ya se tiene el péptido del tamaño
adecuado ese péptido se va a unir a la hendidura y cuando ya lo tenemos unido a
la hendidura ya se tiene la molécula estable que por transporte vesicular va
salir a presentarle un péptido a un LTCD8.
Las
otras chaperonas son estabilizadoras, aquí la Tapasina es la que asegura.

Hay muchos microorganismos que bloquean la
acción de cualquiera de esas moléculas, por eso evitan que sean presentados y
por eso se generan infecciones crónicas. Un microorganismo que anule cualquier
punto, va a evitar la presentación, va evitar su reconocimiento y por lo tanto
van a tener éxito infectando al individuo.
¿Cuál es el tamaño de péptidos más adecuados
para presentar por moléculas de MHC clase I? Es de 9 aminoácidos. El tamaño de 8 a 11, pero los péptidos que generan
respuestas inmunes más fuertes son péptidos de 9 aminoácidos.
Cuando una célula está infectada o se está
transformado para convertirse en una célula tumoral, esta célula va a recibir
el estimulo de la citoquina que se llama INTERFERON g. En una respuesta inmune a virus o a tumores
se genera mucho INTERFERON g. Cuando la célula es estimulada por el
Interferon g, en esa célula se induce la transcripción de dos genes que se llaman
LMP2 y LMP7 que también son genes NO clásicos del MHC, estas dos moléculas se
pueden unir al proteosoma y convierten el proteosoma en INMUNOPROTEOSOMA. La
importancia del Inmunoproteosoma es que va a generar péptidos de 9 aminoácidos,
lo que va a inducir que la acción de los LTCD8 sea más fuerte contra esa célula,
facilitando así la eliminación de las células infectadas o de las células
tumorales. La acción del proteosoma por lo tanto se mejora cuando se le asocia
el LMP2 y el LMP7.
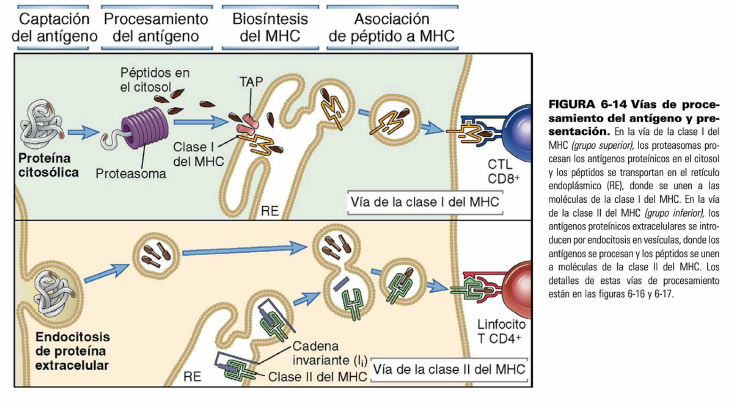
Aquí se puede comparar la via de Clase I y la
de Clase II. En ambas vías se dan los mismos eventos: Ingreso o síntesis de las
proteínas, procesamiento de las proteínas, síntesis de las moleculas, unión de
los péptidos con las moleculas y presentación.
¿Cuál es la finalidad de la presentación de
antígenos?
Cuando se hace presentación de antígenos por
via de clase II el resultado es activar LTCD4 en respuesta a la activación de
los LTCD4 la celula que hizo la presentación va a ser estimulada a mejorar su
acción efectora (se potencia su acción efectora), gracias a las citoquinas que
secretan el LTCD4. Entonces la presentación de péptidos a LTCD4 va a potenciar
la acción efectora de la celula que hizo la presentación.
La finalidad de las celulas que hacen la
presentación de antígenos a los LTCD8 es la inducción de apoptosis en la celula
que hizo la presentación, porque el LTCD8 reconocera esa celula como una celula
tumoral o infectada.
Por esto a los LTCD4 se le llaman ayudadores
o cooperadores, y a los LTCD8 citotoxicos.
apuntes de inmunología medicina - cuarto semestre UL



No hay comentarios:
Publicar un comentario