






MADURACIÓN Y
ACTIVACIÓN DE LINFOCITOS B
A partir de una célula madre aparece un
linfocito, el cual entra en un primer estadio PROLINFOCITO, luego pasaría a
PRELINFOCITO, por último, pasaría al estadio de linfocito INMADURO y que de
aquí saldría hacia al bazo a terminar su proceso de maduración, estos
linfocitos ya maduros se ubican en los órganos linfoides secundarios y se
denominarían linfocitos Vírgenes.
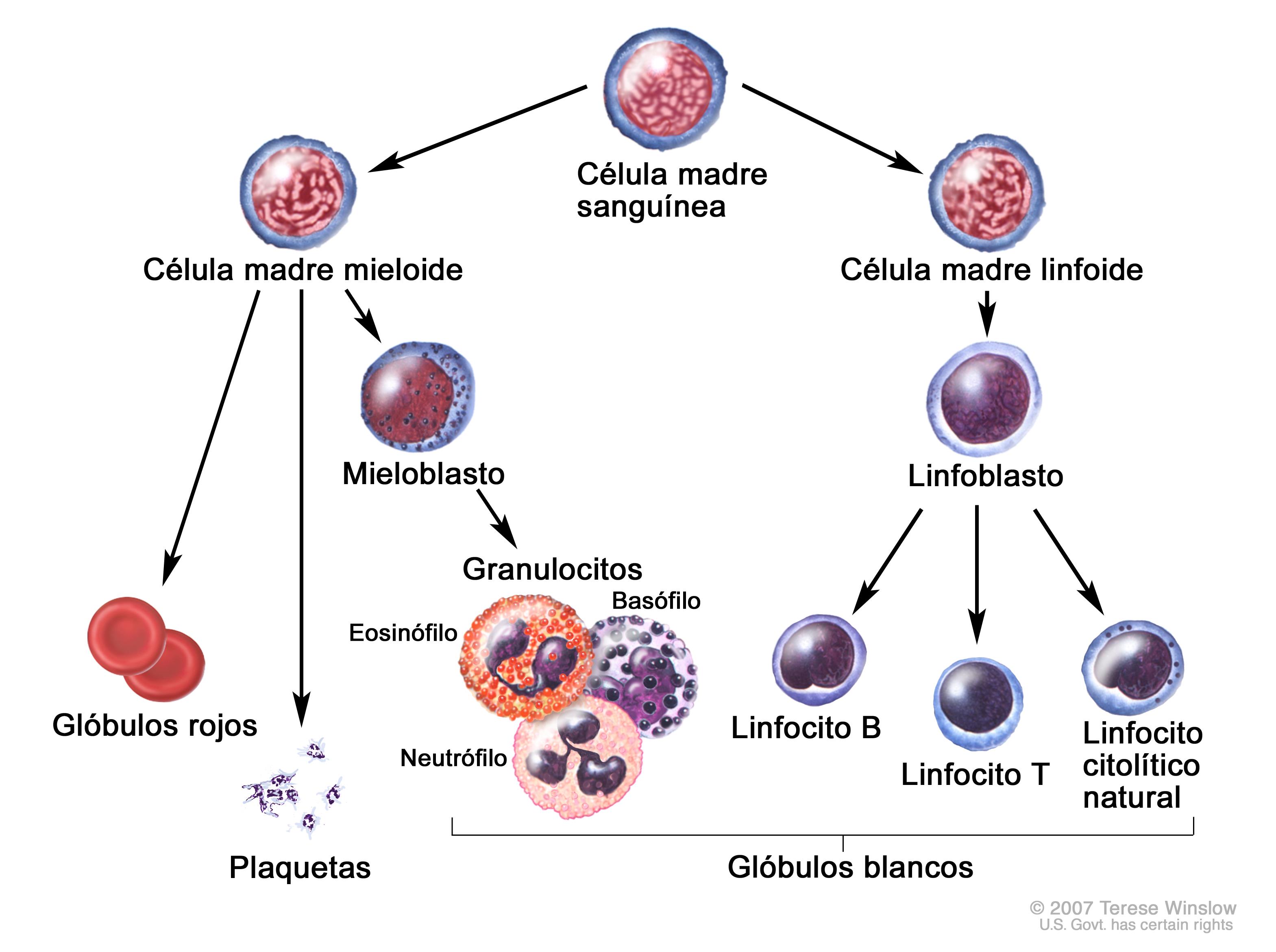
Desde una célula madre, se origina un
linfocito B, el primer estadio es el prolinfocito el cual se multiplica
estratégicamente antes de cambiar de estadio para garantizar que alguno de la
clona pueda llegar al siguiente estadio, para pasar de PRO a PRELINFOCITO, el
linfocito B debe expresar la cadena pesada μ y las cadenas sustitutas, aquellos
que no lo logran por algún motivo si no lo expresa muere por apoptosis, sin
embargo existe 2 oportunidades de expresar esta cadena pesada, si se expresa
normalmente ya sea el alelo de la madre o el alelo del padre, se silencia la
otra cadena del otro padre, no se puede expresar los dos alelos de los padres
al tiempo. El siguiente estadio es el linfocito INMADURO, para que el
PRELINFOCITO pasa a este estadio necesita expresar las CANEDAS LIVIANAS, para
esta cadena tenemos 4 alelos. 2 alelos de cadena KAPPA del padre y de la madre
y 2 alelos de cadena LAMDA del padre y de la madre, si se expresa una cadena
perteneciente a un alelo de algún padre los otros se silencian, una vez que se
expresa la IgM en la membrana el linfocito tendrá que evaluar la autoreactividad
de su BCR, el receptor tuvo que haber aprendido a reconocer lo propio de lo
extraños, la prueba consiste en
colocarle antígenos propios, si el linfocito se une con alta afinidad a un
antígeno propio desencadena una señalización extracelular y genera la
activación de las proteínas PROAPOPTOTICAS, generando un PROCESO DE SELECCIÓN
NEGATIVA, es la lisis o la muerte por apoptosis, si esta unión se realiza con
una afinidad baja el linfocito selecciona POSITIVAMENTE. El linfocito
seleccionado de forma positiva va entonces finalmente hacia los órganos
linfoides secundarios, donde tendrá contacto con antígenos foráneos y una vez
que se activa, proliferara y luego cumplirá la acción efectora, la acción
efectora del linfocito B es generar anticuerpos.
A través de los segmentos génicos que se
encuentran en los cromosomas lográbamos formar la región variable y la región
constante de la cadena pesada, la región variable y la región constante de la
cadena liviana. la unión de la sección V, D, J, se produce al azar, esta unión
al azar no es tan sencilla de entender ya que el linfocito B es una célula que
juega con su ADN, La sección V, la sección D, y la sección J, son segmentos que
se encuentran separados, para que esto se organice o se junte se necesita cortar
por medio de endonucleasas el ADN en sitios que se encuentran señalizados, una
vez se corta en estos sitios quedan más cerca los segmentos , sin embargo aún
no se encuentran totalmente unidos los segmentos, se debe reparar el ADN y para
esto se debe colocar NUCLEOTIDOS P o M. Dependiendo si tenían el molde, los
nucleótidos que se colocan en el espacio dañado eran NUCLEÓTIDOS PALINDROMICOS,
sin embargo si quedaba un hueco o espacio pequeño dentro del ADN se adiciona
NUCLEÓTIDOS M (NUEVOS) , por medio de una enzima llamada Dehoxinucleotidil
Transferasa TDT. Es la enzima encargada de colocar los nucleótidos en esos
pequeños espacios que quedaron, esto sucede de igual manera en el TCR. Aunque
se unan los mismos segmentos de la sección V, D, J repetidas veces, lo
nucleótidos que se adicionan siempre serán diferentes, esto hace que la
especificidad de un anticuerpo sea diferente, haciendo que aumente el
repertorio del BCR, cada uno específico para determinado Epitope. Una vez que
el V, D, J, ya está listo y reparado tendremos el anticuerpo o BCR que necesita
cada linfocito. Mencionamos también que cuando se trata de un BCR, se sintetiza
una región transmembranal e intracitoplasmatica para poder anclarse en la
membrana, sin embargo cuando se trata de un anticuerpo que se va a secretar no
se necesita estas regiones.

Los linfocitos INMADUROS se dirigen
hacia el bazo, terminan el proceso de maduración en este órgano y recirculan
entre los órganos linfoides esperando el antígeno para el cual es específico.
Un linfocito EFECTOR es aquel linfocito que empieza a secretar anticuerpos.
Este proceso de Maduración sucede en la medula ósea, cuando el linfocitos
expresa la IgM en sus membrana sale hacia el bazo y pasa por unos estadios
transicionales, los cuales son estadios que le permiten al linfocito expresar
la IgD convirtiéndose en este caso en linfocitos B2. Una vez el linfocito B
expresa La IgM, sale de la medula ósea y se dirige hacia el bazo, en este
órgano pasa a un estado transicional 1 y transicional 2, este estado transicional
permite a la célula realizar microarreglos para expresar la IgD, únicamente se
produce en los linfocitos B2 Foliculares. Inicialmente se manifestó la
existencia de tres subpoblaciones de linfocitos: Linfocitos B1, se originan en el feto, lo que se reconoce en el
humano es la IgM que es el primer anticuerpo secretado en la etapa fetal, la
IgM reconoce antígenos TIMOINDEPENDIENTES antígenos como carbohidratos,
lípidos, glicolipidos, Acido nucleicos, polisacáridos. Los linfocitos B2 que se
pueden diferenciar en: linfocitos B2
FOLICULAR los cuales poseen IgM e IgD y los linfocitos B2 de la zona Marginal del bazo. El linfocito sale de la
medula ósea una vez se encuentra su IgM expresada y entra al bazo, entra en
transiciones, es decir algunos se convertirán en linfocitos B2 FOLICULARES y
los otros se convertirán en LINFOCITOS B2 DE LA ZONA MARGINAL DEL BAZO, la
diferencia es que uno expresa IgD y el otro no la expresa. Los linfocitos tanto
B1 como los linfocitos B2 tienen la característica que en su membrana tiene IgM
y que producen y secretan cuando se activan IgM, la activación se produce por
el reconocimiento de un antígeno timoindependiente.
Los linfocitos
MZB: tiene
IgM en su membrana Y la proteína CR2, tiene la característica que cuando se
activa por antígenos no proteicos produce IgM.
Los linfocitos
B2 Foliculares:
tienen en su membrana IgM e IgD. La característica de los LB2 foliculares es
que cuando se activa por antígenos proteicos (timodependientes) produce IgM y
produce la reacción del centro germinal, puede cambiar el isotipo, siendo capaz
de producir IgA, IgG e IgE. El cambio de isotipo consiste en que este mismo
linfocitos pueda cambiar la región constante del anticuerpo, y producir
anticuerpos tipo IgA, IgG e IgE, todo esto es posible debido a que el ADN de la
región constante de la cadena pesada NO se corta ni se junta como los segmentos
de la región variable, en el transcrito primario se produce el corte y empalme de
la sección que se está necesitando.
El PRELINFOCITO tiene expresado sobre su
membrana la CADENA PESADA μ y las cadenas sustitas.
La Igα e Igβ, son las proteínas
señalizadoras del receptor, comienzan a sintetizarse en el estadio de
PROLINFOCITO, no se expresan aun sobre la membrana, en el PROLINFOCITO ocurre
los rearreglos para iniciar el proceso de expresión, La Igα
e Igβ son
expresadas en el estadio de PRELINFOCITO,
tienen como
características los Motivos ITAM, los cuales tiene como característica la
capacidad de fosforilar tiene tiroxina Kinasa en su composición y esta proteína
es suceptible a ser fosforilada. Los
receptores no fosforilan, no hace otra cosa que recepcionar el antígeno, los
linfocitos B recepcionan o se unen a antígenos grandes, puede procesarlo y
presentarlo, estos linfocitos reconocen los epitopes para el cual el receptores
es especifico.
El BCR está conformado por IgM (una cadena
pesada μ y una cadena liviana), está conformado por Igα e Igβ
(prerreceptor), este prerreceptor le permite
al linfocito realizar una señalización intracelular de supervivencia o
sobrevivencia, cuando se expresa la cadena μ de algún padre se silencia la
expresión de la otra cada derivada del otro alelo paterno, gracias a un proceso
de señalización que produce el prerreceptor para el silenciamiento del otro
alelo paterno, debido a que si expresamos un alelo paterno y un alelo materno
tendríamos especificidad a cualquier cosa y no justamente al epitope del
antigeno. Los linfocitos B tienen como característica que son específicos a
cierto epitope, una vez que se expresa el alelo paterno adecuado se silencia
por señalización el otro y el prelinfocito prolifera para pasar a la siguiente
prueba que lo llevara al siguiente estadio. Para que se produzca la mitosis
debe existir una señalización intracelular en el prelinfocito. Se estimula la recombinación genética de las
cadenas liviana, necesitamos las cadenas livianas que estabilizan la cadena
pesada. Las cadenas sustitutas que cumplieron la función de cadenas livianas se
endocitan y son degradadas. La Igα e Igβ después
de su expresión sobre la membrana siempre van acompañar el linfocito B, debido
a que son las proteínas encargadas de la señalización intracelular así como lo
son la cadena z y el CD3 en el linfocito T.
Unos
autores en el 2010 dijeron que la comunicación está dada por unos aminoácidos
que se encuentran en el prereceptor que están cargados positivamente y que se
unen a las células estromales de la medula ósea con aminoácidos negativos, esta
unión negativo – positivo, hace que se señalice para que suceda todo lo
mencionado, rearreglo de la cadena liviana, silenciamiento de alelo del padre
que no expreso primero las cadenas, endocitosis de cadenas sustitutas para que
salga la IgM completa (unión de cadena pesada con cadena liviana), esta IgM
completa se encuentra en un linfocito inmaduro, aun no se conoce que hace que
el prereceptor se active en estos estadios para generar la señalización
intracelular y se genere los procesos determinados de cada estadio.

La función de los PRERRECEPTORES Igα e
Igβ SON:
- · Silenciamiento del alelo paterno que no expreso de primero la cadena Pesada.
- · Proliferación de PRELINFOCITOS B
- · Estimulación de la recombinación de los segmentos o secciones de la cadena liviana.
- · inhibición de transcripción de la cadena Liviana sustituta
Una vez que hay una
interacción de los prerreceptores sucede lo nombrado anteriormente en los
PRELINFOCITOS B.
El señor Odger Bruton un pediatra militar de la década de
los 50 observo paciente con infecciones
a repetición, en esta década no se conocía las inmunodeficiencias, el doctor bruton
tomo muestras de sangre de estos pacientes, quería comprobar los niveles de
anticuerpos secretados por estas
personas y observo que no secretaban
grandes cantidades de anticuerpos su secreción casi era nula, la enfermedad
que el descubrió se denomina Agammaglobulinemia, los anticuerpos cuando son
medidos en sangre, pertenecen a la fraccion GAMMA de las proteínas que hay en sangre, las
inmunoglobulinas también son denominadas Gammaglobulinas, por esta razón se
denomina A – GAMMA globulina (sin inmunoglobulinas) enfermedad ligada al cromosoma
X porque se vio solamente en niños y al hacer un estudio de ¿por qué estos
niños no producían anticuerpos? se dio cuenta que en el PRELINFOCITO B la proteínas kinasa de bruton está funcionando y
que aquellos niños que tienen mutación en esta kinasa, sus linfocitos maduran
hasta los PRELINFOCITOS B. un PRELINFOCITO no puede secretar inmunoglobulinas
ya que no ha cumplido su proceso de maduración. A estos niños se les mide los
linfocitos B maduro y son muy mínimos (quizás son los linfocitos B1 fetales que
pueden secretar IgM pero no cambiar de isotipo). Los linfocitos B2 son
producidos hasta los PRELINFOCITOS, es por esto que los niveles de IgM son
bajos.

Parece que los linfocitos B1
permanecen vivos por algún mecanismo en específico.
A esta tiroxina kinasa le
dieron el nombre de bruton en honor al médico que descubrió esta patología,
esta kinasa no es solamente importante en el proceso de maduración del
linfocito b sino que también en el proceso de activación y señalización.
Cuando el linfocito B maduro
encuentra su antígeno determinado, se activa, se multiplica y la mayoría de la
clonación se diferencia a células plasmáticas productoras de anticuerpo y
alguno se guardan como células de memoria, las células de memoria sufren un
proceso que se llama la reacción del centro germinal, este linfocito B será un
mejor linfocito dispuesto para la defensa debido a que ha sido un linfocito
maduración de la afinidad y cambio de isotipo. La maduración de la afinidad
consiste en el aumento la fuerza con que se une el receptor del linfocito b al antígeno,
la segunda vez que se infecta una persona. El cambio de isotipo consiste en que
el linfocito B ya no producirá IgM sino que inducirá la producción de la
inmunoglobulina adecuada o especializada para ese microorganismo, si hay un
microorganismo en la mucosa no necesitara pasar por IgM sino que secretara
directamente IgA, si necesita Ig para un helminto se secretara directamente
IgE, si se necesitan Ig para neutralizar un virus se produce IgG, esto
garantiza que sea mucho más rápido y efectiva la respuesta secundaria,
normalmente cuando un linfocito se activa, puede producir hasta 5000 células
plasmáticas, que producirán 10^12 anticuerpos por día.
Los linfocitos se empiezan a madurar en la
medula osea, salen como linfocitos inmaduros hacia el baso pasan por un estado
transicional y terminan de madurar en este órganos, luego comienzan a
recircular en todos los órganos linfoides secundarios
Tomaremos como ejemplo el
ganglio, en la corteza del ganglio encontramos unos espacios, que son los
folículos. Resulta que los linfocitos B siempre se van a ubicar en los
folículos, que en este momento se denominaran los folículos primarios, ¿por qué
los linfocitos se ubican específicamente en este espacio? debido a las
quemoquinas y receptores de quemoquinas, para que el linfocito B migre a este folículo
va a expresar el CXCR5 el cual es atraído por CXCL13, el folículo primario es
rico en CXCL13. Los linfocitos T vírgenes se ubican en la paracorteza del
ganglio linfático, esta migración es igual a la anterior, debe expresarse un
receptor de quemoquinas CCR7 y las quemoquinas que abundan en este espacio son
la CC19 y CCL21. Los linfocitos B pueden llegar por los vasos linfáticos
aferentes o por el torrente sanguíneo, En la medula del ganglio se van a ubicar
los linfocitos que van de salida, ya sea Vírgenes, activados u efectores van a
salir por los vasos sanguíneos o por los vasos linfáticos eferentes.

En el foliculo primario del
nodulo linfático se presentan unas celulas dendríticas foliculares, son celulas
mostradoras, NO SON CELULAS PRESENTADORAS DE ANTIGENO, son las únicas celulas
que no procesan y presentan el antigeno si no que toman el antigeno con sus
receptores y los muestran. en lo vasos aferentes no tan solo llega células
presentadoras de antígenos proveniente de la infección o lesión, sino también
los microrganismo y los antígenos que son recogidos por la linfa. Estas células
y moléculas van a entrar, la célula dendrítica va entrar a la paracorteza para
tratar de activar al linfocito T virgen, y el antígeno va tratar de llegar al
linfocito B que se encuentra en el folículo.
De los vasos aferentes llegan
los antígenos a la corteza, si el antígeno es pequeño se sabe que por los
ductos que hay en el ganglio, pueden llegar directamente y entrar hacia el folículo
donde se encuentra todos los linfocitos B inactivos. En la corteza,
específicamente en folículos se encuentra las células dendríticas foliculares y
los Linfocitos B vírgenes. ¿Como llega el antígeno al linfocito B?
Algunos científicos hicieron
experimentos donde marcaron los antígenos, los inyectaron y le siguieron el
recorrido, y se dieron cuenta que los antígenos más pequeños podrían entrar
directamente al folículo donde se encentran los linfocitos B vírgenes por un
ducto que lleva directamente al folículo primario. El BCR, puede reconocer
cualquier naturaleza química, del antígeno. Puede entrar todo microorganismo y
solo se debe reconocer el epitope de este. Aquellos antígenos que eran grandes,
no podían entrar por el ducto, eran transportados por macrófagos llamados
MACROFAGOS DEL SENO CAPSULAR, y CÉLULAS DENDRÍTICAS DIFERENTES que forman una
malla y pasaban el antígeno específico para el linfocito B. El macrófago
también podía viajar y entregárselo al linfocito B específico para este
antígeno, El macrófago del seno subscapular, su prioridad no es fagocitar,
procesar y presentar, ellos toman antígenos y los transportan, los pasa por
cada linfocito hasta que se une con un linfocito con gran afinidad para este
antígeno.
Hay unas células dendríticas
que también están involucradas en transportar este antígeno, hacia el folículo
primario. Los macrófagos no se encargan de modular la afinidad tan solo son
transportadores. Cuando el antígeno llega al linfocito B virgen, este linfocito
se activa pero generalmente necesita otros estímulos, como las citoquinas
secretadas por linfocitos T. Cuando llega el estímulo, los linfocitos B se
multiplican hace expansión clonal y se diferencia en una célula efectora o
célula plasmática productora de IgM. Si se saca sangre a una persona que
recientemente ha tenido una infección, encontraremos muchas cantidades de IgM
durante de los primeros 8 días.
La mayoría de las clonas se
diferencian a células efectoras productoras de anticuerpos, tan solo unas se
devolverán al folículo y realizaran la reacción del centro germinal. Las clonas
se diferenciaran en células plasmáticas de corta vida son aquellas que están
secretando IgM. Otras se devolverán al folículo y producirán la reacción del
centro germinal. Lo que sucede en la reacción del centro germinal es: MADURACIÓN DE LA AFINIDAD, CAMBIO DE
ISOTIPO, se GENERARAN CÉLULAS PLASMÁTICAS DE LARGA VIDA Y CÉLULAS DE MEMORIA. Únicamente
puede realizarse por los Linfocitos B2 FOLICULARES.
El linfocito B se activa ante la unión
con el antígeno específico, únicamente con el BCR se puede activar, recordemos
que los linfocitos B son células presentadoras de antígenos ósea que puede
entocitar, procesar y presentar los péptidos derivados de los antígenos. El
linfocito B reconoce otro epitope del antígeno presentado por las moléculas del
MHC clase 2 a los linfocitos T. No necesariamente tiene que ser el mismo
epitope que se presente al linfocito T para activar los linfocitos B, ya que pueden
ser epitopes diferentes. El receptor del linfocito B (BCR) está acompañado de
un correceptor, el cual está conformado por: TAPA- 1/CD81, CD19, CR2 O CR21. El linfocito se puede activar
únicamente con el receptor y el correceptor. Sin embargo en algunas ocasiones
al microorganismo se le queda unido el C3d (un producto del clivaje de C3, este
tenía una cadena α y una cadena β, en el momento de proteolizar, la fracción
más pequeña se denominaba C3a, esta se separaba del complejo y lo que quedaba
se unía covalentemente a la membrana del microorganismo a este se llamó C3b,
sin embargo para protegerse del C3b debido a que este se encarga de opsonizar y
de activar la vía alternativa del complemento los microorganismos expresan
proteínas enzimáticas que proteolizan este C3b, en C3c y C3d, por esto algunas
veces queda pegado el C3d al microorganismo). Los linfocitos B puede reconocer
este C3d que se ha quedado pegado al microorganismo a través de su correceptor,
induciendo una activación con mucha más fuerza del linfocito B por medio de su
correceptor, si este microorganismo no tiene el C3d no pasa absolutamente nada
ya que de igual forma se activara el linfocito B. Si al microorganismo se
encuentra pegado el C3d el CR2 -receptor 2 del complemento, lo reconoce pero
quien señaliza es el CD19 ya que contiene motivos ITAM. Recordando que el
prerreceptor se encarga de señalizar la unión del BCR con el antígeno, mientras
que el CD19 señaliza la unión del C3d con el CR2. Cuando el BCR y el CORREEPTOR
se unen con sus determinados ligando las fosforilaciones son mucho más rápidas,
más potentes y hay activación es rápida.
Los TLR en aquellos
linfocitos que no tienen correceptor, pueden actuar en este mismo sentido,
cuando se reconoce PAMPS de microorganismo se induce una señalización, estos
PAMPS van a ser reconocido más por los LINFOCITOS DE LA ZONA MARGINAL DEL BAZO
y los LB1, los TLR pueden actuar como correceptor y ayudar a esta señalización.
Los LB1 no contienen el Correceptor.
El BCR una vez se une con el antígeno,
tiene un cambio conformacional. La IgM que es el receptor del linfocito amplía
su tamaño y hace que las proteínas tirosinas Kinasas (PROTEINAS DE LA FAMILIA
SRC) por ejemplo: proteínas LYN, FYN, BIK se pegen. La balsa lipídica es un
dominio, hará que todos los receptores y las moléculas necesarias se junten
para que puedan hacer una sinapsis inmunológicas, Todos los componentes del BCR
se juntan (Igα e Igβ, IgM e IgD) y se pueda producir una señalización. La
quinasa LYN que está pegada a la membrana empieza a fosforilar los motivos ITAM
de la Igα e Igβ, CD19 que es el señalizador del correceptor contiene motivos
ITAM que fosforilan, se observa que una vez fosforilados los motivos ITAM DE LA
igα e igβ atra una tirosina kinasa llamada SYK e induce la fosforilacion de
PI3-kinasa y se continua la señalización.
Las balsas lipídicas unen los
BCR de la superficie de los linfocitos, y para que se active el antígeno
realmente debe hacer una reacción cruzada entre las dos inmunoglobulinas, las
regiones variables de las inmunoglobulinas deben tener contacto con el
antígeno.

La parte rosada de la
diapositiva son las balsas lipídicas, se deben unir los receptores,
correceptores y prerreceptores para que haya una activación de LYN (tirosinas
quinasas que está cerca de la membrana en la balsa lipídica) y fosforilacion de
BLINK y activación de SYNK.
LYN es una de las tiroxinas
quinasas que va a fosforilar Los motivos ITAM de las igα e igβ, a su vez atrae
a SIK, la proteínas SIK atrae a SLP-65 que es una proteína compendiadora,
unidora de otras. La proteína Kinasa de Bruton va activar la proteína
FOSFOLIPASA C-GAMMA 2, toma los fosfolípidos de la membrana, el
fosfoinositoldifosfato y lo convierten en diacil-glicerol y inositol
trifosfato, es igual en todas las células. El diacil glicerol, activa la
proteína Kinasa C, que induce la activación del factor nuclear TAPA B.
El inositol trifosfato es una
molecula que extrae el calcio donde se encuentre guardado en la celula, y
permite que forme poros para que entre calcio del exterior celular, esto induce
la activación de unas enzimas dependiente de calcio como la calcineurina y la
calmodulina, estas enzimas van activar el factor nuclear del linfocito B –
NFAT. Por otro lado la vía de las RAC quinasas va activar los factores ERK Y
JNK, para que se produzca el factor nuclear AP-1. Es igual que la señalización
de linfocitos T. La activación del linfocito B sirve para activar los factores
nucleares, los cuales se introducen en el ADN y generan la síntesis de
ANTICUERPOS.
Los linfocitos B son también
células presentadoras de antígenos, producen una endocitosis con ayuda de
CLATRINA (molecula que ayuda a formar los endosomas), genera la endocitosis de
los antígenos con los BCR y cuando está en el estadio de endosoma temprano hay
una señalización que separa el BCR del antígeno, en dos vesículas, una se
transforma en una vesícula reciclante que vuelve y saca el BCR, y la otra
vesícula se vuelve en un endosoma tardío, en este endosoma tardío es donde el
antígeno va a tener contacto con EL MHC de clase 2 que viene con su CLIP y cadena invariante y una vez que se
encuentran, es donde ocurre la degradación de la cadena invariante, la quitada
del CLIP por el HLA-DM y la unión de uno de los péptidos a la molécula MHC
clase dos para que luego se pueda expresar, para una presentación antigénica en
el contexto del MHC clase 2. Lo proteico del antígeno es aquí, donde se va a
visualizar para el linfocito T
Un linfocito B se tiene que
activar para poder producir inmunoglobulinas M y luego producir una reacción
del centro germinal, cuando los antígenos son timodependientes (esto quiere
decir que son antigenos proteicos que son suceptibles a ser presentados a los
linfocitos T), después de la activación del linfocito B y del linfocito T, debe
haber una interacción de linfocitos T con antigeno, ellos deben encontrarse
geográficamente en paracorteza y corteza. Hay una proliferación y
diferenciación de linfocitos b que genera una secreción de anticuerpos y
posteriormente una reacción del centro germinal.
La reacción del centro
germinal: 1) un linfocitos B virgen se activa ante el microorganismo completo
reconociendo el epitope, cuando se produce la señalización antes mencionada se
activa proteínas antiapoptoticas, para que sobrevida e induce una gran
multiplicación o expansión clonal.
2) empieza a expresar
moléculas coestimuladoras necesarias para esta sinapsis inmunológica. Se
incrementa la expresión de receptores de citoquinas, los linfocitos T secretan
citoquinas para ayudarle al linfocito B a activarse mejor, para que pueda
producir una señalización importante para que cumpla con su función, para que
el LB virgen se ubique en el folículo
debe expresar un receptor CXCR5 para unirse con su ligando CXCL13, sin
embargo cuando son activados por los antígenos que se introdujeron al
folículo por los ductos o por transporte mediado por macrófagos comienza a
endocitar, procesar y expresan sobre su membrana un MHC de clase 2 con el
péptido para presentarlo al LT , comienzan a expresar CCR7 que los induce a
migrar a la paracorteza ya que esta es rica de CCL21 y presentar el antigeno al
linfocitos T que se encuentra en la corteza, las células dendríticas que
endocitaron, procesaron el antigeno y unieron un péptido de este a su MHC de
clase 2 se dirigen a los nódulos linfáticos, buscaron el TCR para el cual es
especifico el antigeno, una vez se une con el TCR y es estimulado por las células
coestimuladoras, se activa y se multiplica, alguno de los clones se diferencian
en linfocitos TH1, linfocitos TH2 y linfocitos TH17 pero existen otras clonas
de LT que empiezan a expresar CXCR5 sobre su membrana, los linfocitos B
anteriormente mencionados, empiezan a expresar CCR7 que induce su migración
hacia la paracorteza, ahora estos linfocitos T que han expresado CXCR5 inducen
su migración hacia el folículo primario y poco a poco las dos clases de células
se van acercando justo en la salida del folículo, los linfocitos B presentan el epitope a los linfocitos T, se
genera una interacción inicial de LT –LB, en esta cooperación los linfocitos T
le secretan citoquinas a los linfocitos B y estos se transforman en células
plasmáticos de corta vida, dura aproximadamente 5 días secretando muchas cantidades
de IgM, Son estos los anticuerpos que nos van a defender en una infección
inicial o primaria.
Bibliografia: Abul K. Abbas, Andrew H. H. Lichtman, Shiv Pillai, Inmunología celular y molecular. edición 7. Elsevier España, 2012. Graficas relacionadas con post e información
apuntes de inmunología medicina - cuarto semestre UL



No hay comentarios:
Publicar un comentario